Role of termites and fungus in carbon release from tropical forest deadwood
The study examines the differential recovery of fungal and termite functions in deadwood decomposition following rainforest regeneration in Australia. The researchers found that while fungi consistently contribute to carbon and nutrient cycling, termite species richness, composition, and functional roles are significantly reduced in secondary tropical rainforests compared to old-growth forests. This suggests that fungal communities are more resilient to disturbance than termites, with potential repercussions for carbon transfer to soil and the atmosphere across global ecosystems. The study challenges common assumptions about the parallel recovery of fungal and termite functions post-regeneration, emphasizing the need for future research to explore the broader ecological implications for carbon dynamics worldwide.
Citation: The Following Is An Unpublished ‘News And Views’ Style Discussion Written By Me About “Wijas, B. J.; Flores‐Moreno, H.; Allison, S. D.; Cernusak, L. A.; Cheesman, A. W.; Eggleton, P.; Kooyman, R. M.; Powell, J. R. and Zanne, A. E. (2025). Decadal recovery of fungal but not termite deadwood decay in tropical rainforest, Journal of Applied Ecology. https://doi.org/10.1111/1365-2664.70053” for general audience.
Contents
- TITLE:
- KEYWORDS:
- SUMMARY
- INTRODUCTION
4.1 The Interplay of Land-Use Change and Carbon Emissions
4.2 Drivers of Carbon Emissions from Forests
4.3 Composition and Dynamics of Carbon Storage in Forests
4.4 Old Forests as Net Carbon Sinks and Secondary Forests as Initial Carbon Sources: The Role of Deadwood Decay
4.5 Decomposition Processes in Forest Ecosystems
4.6 Fungal and Termite Roles in Wood Decay
4.7 Symbiotic Relationships in Wood Decay
4.8 The Dual Role of Deadwood Decay - Research Design And Evaluation (aka Philosophy of Science or Scientific Method In Action)
5.1 Observations, Concepts and Evidence
5.2 Hypothesis (making a calculated guess about cause and effect)
5.3 Experiment (establishing a system to collect data to evaluate if the guess of cause and effect is factually true or not)
5.4 Experimental Methods (experimental setup and data collection from it)
5.5 Experimental Results (results are obtained by doing analysis on the data)
5.6 Inference (taking results into account, hypothesis is accepted or rejected): - METHODS
6.1 Study Overview
6.2 Canopy Structure Differences
6.3 Canopy Openness
6.4 Diversity and Complexity
6.5 Changes by 2018
6.6 Termite and Fungal Community Study
6.7 Methodological Differences
6.8 Decay Process Recovery Assessment
6.9 Experimental Setup
6.10 Collection and Analysis of Wood Blocks
6.11 Fungal Community Analysis - RESULTS
7.1 Figure 1 of Paper: Decay of Termite-Discovered Deadwood in Secondary vs. Old Growth Rainforests
7.2 Figure 2 of Paper: Fungal Decay of Deadwood and Community Composition - DISCUSSION
8.1 Decay Processes in Rainforest Ecosystems
8.2 Interactions Between Termites and Fungi
8.3 Nutrient Recycling and Ecosystem Health
8.4 The Ecological Significance of Deadwood
8.5 Role of Fungi and Statistical Analysis in Decay Rates
8.6 Implications of Disturbances on Decay Processes
8.7 Exploring the Influence of Evolutionary History on Deadwood Decay Across Global Forest Ecosystems
8.8 Alexander von Humboldt and The Ecological Network: From Microbial Decomposers to Global Carbon Cycles
1. TITLE:
Role of termites and fungus in release of carbon from tropical forest deadwood to carbon nutrients in soil and carbon emission in air.
2. KEYWORDS:
decadal recovery , fungal decay, termite deadwood, tropical rainforest, deadwood decomposition, rainforest regeneration, fungal communities, carbon cycling, nutrient cycling, ecological implications.
3. SUMMARY
Decadal Recovery of Fungal but Not Termite Deadwood Decay in Tropical Rainforest” by Baptiste J. Wijas et al. offers valuable initial insights into the differential recovery of fungal and termite functions in deadwood decomposition following rainforest regeneration in Australia. The researchers designed the experiment, secured funding, and analyzed the collected data to reveal that while fungi contribute consistently to carbon and nutrient cycling, termite species richness, composition, and functional roles are significantly reduced in secondary tropical rainforests compared to old-growth forests. This indicates that fungal communities are more resilient to disturbance than termites, with potential repercussions for carbon transfer to soil and the atmosphere across global ecosystems. Notably, the authors challenge common assumptions about the parallel recovery of fungal and termite functions post-regeneration, emphasizing the need for future research to explore the broader ecological implications for carbon dynamics worldwide.
4. INTRODUCTION
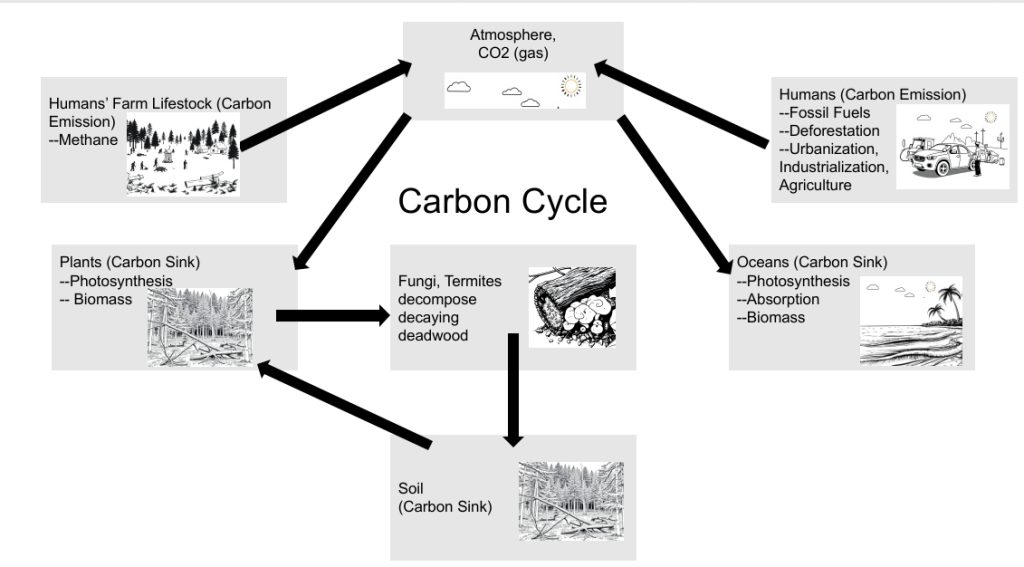

4.1 The Interplay of Land-Use Change and Carbon Emissions The interplay between land-use change, forest decay processes, and global carbon emissions is a critical aspect of current environmental discussions, particularly as we confront the realities of climate change. Deforestation and forest degradation have emerged as significant contributors to anthropogenic carbon dioxide (CO₂) emissions, releasing carbon that is sequestered within trees, vegetation, and soils into the atmosphere. While fossil fuel combustion remains the predominant source of global CO₂ emissions, the role of land-use change in the carbon cycle and its contribution to greenhouse gas accumulation cannot be overlooked. According to the Global Carbon Project’s 2023 report, which assesses carbon budgets with data extending through 2022, land-use change was responsible for approximately 4.2 gigatonnes of CO₂ (GtCO₂) emissions in that year alone. When compared to total CO₂ emissions from fossil fuels and industrial processes, which peaked at 41.0 GtCO₂ in 2022, it becomes evident that land-use changes accounted for roughly 10.24% of all anthropogenic emissions, highlighting a substantial impact on climate change dynamics, while the remaining 90% primarily resulted from the combustion of fossil fuels.
4.2 Drivers of Carbon Emissions from Forests To better understand this issue, it is essential to investigate the primary drivers behind carbon emissions from forests and their cascading consequences. Healthier forests can function as robust carbon sinks, effectively absorbing more CO₂ than they release. However, when these forests undergo deforestation, forest degradation, or are exacerbated by wildfires, they can transition into significant sources of carbon emissions. Deforestation, which involves the permanent clearance of forests for agriculture—such as cattle ranching, soy harvesting, or palm oil plantations—alongside infrastructure expansion, mining activities, and urbanization, is one of the leading causes of forest carbon emissions. In these cases, the carbon stored within biomass and soil is released into the atmosphere. Forest degradation, characterized by practices like selective logging and excessive fuelwood removal, also plays a vital role. Such degradation reduces the density of forests and their carbon storage capacity, making them more vulnerable to fires. Natural and human-induced fires contribute to significant carbon emissions, particularly when compounded by climate change and land-clearing practices. Forest wildfires have increased in frequency and destructive force due to intentional fires started by humans to clear forest land and due to climate change. Additionally, the destruction of peatlands, rich in ancient carbon stores, can release vast amounts of CO₂, especially when these areas are drained or burned for agriculture.
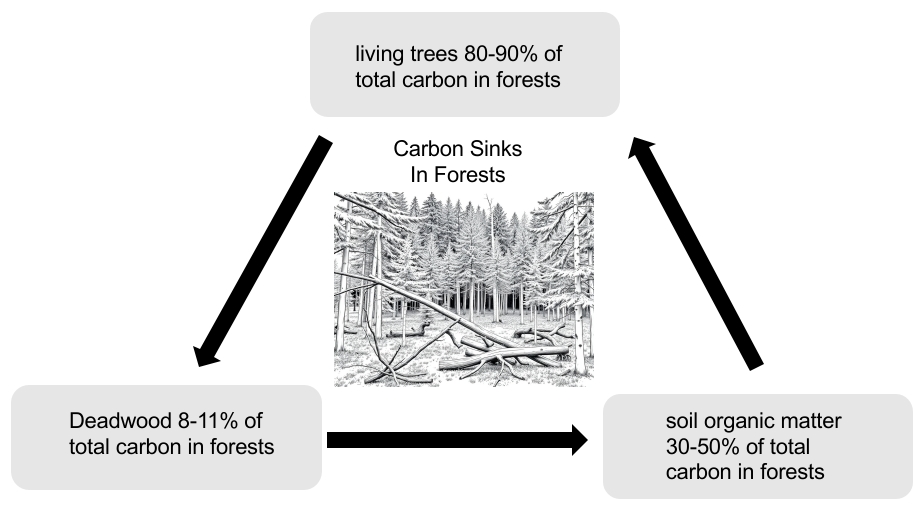
4.3 Composition and Dynamics of Carbon Storage in Forests Understanding the composition and dynamics of carbon storage within forests is crucial. Live trees account for a significant portion—approximately 80-90%—of the total carbon stock, primarily storing carbon through photosynthesis in trunks, branches, leaves, and roots. Beneath this, soil organic matter represents another 30-50%, comprising decomposed plant and animal materials that play a critical role in long-term carbon sequestration. Other important components of forest carbon storage include understory vegetation and the litter layer, both contributing smaller amounts but remaining essential in nutrient cycling processes. Deadwood, which makes up about 8% of total carbon stocks, is also significant in the context of carbon cycling, as its decomposition is mediated by various organisms, notably fungi and termites.
4.4 Old Forests as Net Carbon Sinks and Secondary Forests as Initial Carbon Sources: The Role of Deadwood Decay
Old-growth forests typically serve as net carbon sinks, absorbing more carbon than they release, even when considering the carbon emitted during the decay of deadwood. These ancient ecosystems store substantial amounts of carbon in both living biomass and deadwood. Trees capture carbon dioxide from the atmosphere through photosynthesis, transforming it into sugars that are then utilized to form wood, leaves, roots, and other plant tissues. This carbon becomes incorporated into the living biomass of the forest—found in trunks, branches, and roots—as well as in deadwood, leaf litter, and the soil. While the decomposition of deadwood does return some carbon to the atmosphere and soil, the ongoing growth and accumulation of organic matter in old-growth forests generally outweigh these emissions. Recent research suggests that most forests older than 180 years continue to sequester carbon, challenging the previous belief that old-growth forests are carbon neutral. In contrast, secondary forests—those that are younger and often managed or recovering from disturbances—often act as net carbon emitters in the initial decades following events like clear-cutting or natural dieback. During this early phase, carbon losses from soil respiration and deadwood decomposition surpass the amount of carbon absorbed by new tree growth. However, as these secondary forests mature over a span of 20 to 30 years, their biomass and leaf area increase, allowing them to transition into net carbon sinks. At this point, the rate of carbon sequestration exceeds emissions, leading to a renewed accumulation of carbon. Deadwood plays a crucial role in the carbon cycle for both types of forests. While old-growth forests harbor more deadwood—which releases carbon during decomposition—they also maintain more stable carbon pools and consistent carbon uptake. In contrast, secondary forests have less deadwood but experience elevated carbon emissions immediately after disturbances. Overall, old-growth forests consistently function as net carbon absorbers, while secondary forests evolve from being net emitters in their early stages to becoming net absorbers as they develop.
4.5 Decomposition Processes in Forest Ecosystems Recent studies indicate notable differences in decomposition processes between old-growth and secondary forests. Specifically, leaf litter in secondary forests tends to decay at a slower rate compared to that in old-growth forests. This deceleration disrupts established nutrient cycling and impairs carbon sequestration efforts, limiting the replenishment of nutrients in the soil and hindering the recovery and resilience of these forest ecosystems. This is particularly alarming given that tropical rainforests serve as crucial global carbon sinks; compromised decomposition processes may significantly decrease their capacity for climate mitigation.
4.6 Fungal and Termite Roles in Wood Decay The decomposition of wood, a key process in carbon release, involves intricate interactions between fungi and termites. Fungi play an essential role by enzymatically breaking down lignin and cellulose, altering the wood’s structure, and facilitating further decay. Interestingly, fungal communities in secondary forests exhibit a faster recovery trajectory that approaches the decay rates observed in old-growth systems over time. Termites also contribute to wood decomposition by physically consuming wood, although their efficiency appears diminished in secondary forests, despite their ability to locate deadwood remaining similar across different forest types. The activity of these organisms is heavily influenced by factors such as forest age and prevailing environmental conditions.
4.7 Symbiotic Relationships in Wood Decay One particularly fascinating aspect of wood decay involves the symbiotic relationship between certain termites and specific fungi. This relationship highlights the complex interactions within forest ecosystems that significantly affect carbon dynamics. Understanding these relationships can further illuminate the intricate balance between carbon storage and emissions in various forest types, offering insights for effective conservation and climate mitigation strategies.
4.8 The Dual Role of Deadwood Decay Deadwood decay plays a critical yet paradoxical role in forest carbon dynamics, acting as both a source of CO₂ emissions and a contributor to soil health. As deadwood decomposes, it releases significant amounts of CO₂ into the atmosphere, which contributes to greenhouse gas emissions. This process involves a variety of organisms, notably fungi and insects, that break down the wood’s complex organic compounds. While this release of CO₂ is a concern in the context of climate change, it is important to recognize that deadwood decay is also essential for returning carbon to the soil. When deadwood breaks down, it enriches the soil with organic matter, improving its nutrient content and structure. This enriched soil can then support new plant growth and promote a healthy ecosystem, as the nutrients released during decomposition are taken up by living organisms. Thus, despite its role in emitting CO₂, deadwood decay essentially serves as a necessary evil; it is integral to the nutrient cycling processes that rejuvenate forests and sustain a wide range of life forms. The balance between the immediate carbon emissions from deadwood decomposition and the long-term benefits to soil health and forest regeneration underscores the complexity of forest ecosystems and the need for thoughtful management practices that account for both carbon sequestration and ecosystem vitality.
5. Research Design And Evaluation (aka Philosophy of Science or Scientific Method In Action)
The overall scientific method involves forming a hypothesis, designing an experiment to test it, collecting and analyzing the data, and then drawing conclusions based on the results. The scientific method starts with observation and collecting data, drawing on pre-existing knowledge and facts. Researchers collect facts about the components of their subject matter and establish logical relations between them. As patterns and relationships are noticed, a hypothesis is generated, which is a testable and falsifiable explanation that can be proven or disproven through experimentation and data analysis. A good hypothesis is based on evidence, specific, testable, and consistent with existing knowledge. In the present study, here are the pair of components and their logical relationships:
5.1 Observations, Concepts and Evidence
1. Deforestation and Carbon Emissions: The removal of forests (deforestation) increases carbon emissions by reducing the amount of carbon that forests can store. Land-use change, which refers to the conversion of land from forested to non-forested areas, primarily occurs through human activities such as agricultural expansion, mining, and urban development. Conversely, reforestation—the deliberate replanting of trees—occurs through sustainable agricultural practices and conservation policies. Carbon emissions, specifically carbon dioxide (CO₂), are one component of the broader category of greenhouse gases, which also includes methane (CH₄), nitrous oxide (N₂O), and synthetic gases like hydrofluorocarbons (HFCs). These gases contribute to climate change by trapping heat in the Earth’s atmosphere.
2. Carbon Cycle Dynamics: Human activities, particularly the burning of fossil fuels (coal, oil, natural gas), deforestation, and industrial processes, release significant quantities of carbon dioxide into the atmosphere. This increase in atmospheric carbon dioxide enhances the greenhouse effect, a natural process that becomes problematic when intensified, leading to global warming and subsequent climate change. Forest wildfires quickly emit lots of CO2, when they are started by humans to clear forest land and due to climate change. Carbon sinks, which are natural or artificial reservoirs that absorb more carbon than they release, include forests, oceans, and soil. These sinks help mitigate the impact of greenhouse gas emissions by removing carbon dioxide from the atmosphere.
3. Forests as Carbon Reservoirs: Healthy forests serve as effective absorbers of carbon dioxide, helping to reduce the impact of carbon emissions. According to the Global Carbon Project’s 2023 report, the disruption of forests contributed 4.2 gigatons of carbon dioxide (GtCO₂), approximately 10% of the total 41.0 GtCO₂ global emissions in 2022. The remaining 90% primarily resulted from the combustion of fossil fuels.
4. Role of Deadwood: Deadwood, which consists of fallen trees and branches, contributes to carbon storage in forests and plays an essential role in nutrient cycling through its decomposition process. Despite carbon release from deadwood decay, continuous photosynthesis (conversion fo CO2 to sugars), continuous growth and organic matter accumulation generally outweigh these emissions. Most forests older than 180 years continue to sequester carbon, undermining the idea that they are carbon neutral. In contrast, secondary forests initially act as net carbon emitters after disturbances but transition to net carbon sinks over 20-30 years as they mature.
5. Decomposition Processes: Organisms such as fungi and termites facilitate the breakdown of deadwood, which releases carbon dioxide and methane as byproducts into the atmosphere. Deadwood constitutes 8-11% of the total carbon stored in forests (the forest carbon sink), with living trees (80-90%), soil organic matter (30-50%), understory vegetation, and forest floor litter comprising the remainder.
6. Secondary Forests and Ecosystem Recovery: Secondary forests, which grow after the original forest has been cleared or significantly altered by human activity, climate change or forest wildfires, differ structurally from old-growth forests (mature, undisturbed forests) but are crucial for ecosystem recovery following human disturbance. Reforestation efforts, including planting native species and restoring degraded lands, promote the growth of these secondary forests, which help mitigate the impacts of land-use change and facilitate ecosystem recovery.
7. Decomposition Challenges in Secondary Forests: Scientific research indicates that leaf litter in secondary growth forests decomposes more slowly than in old-growth forests. This slower decomposition can disrupt nutrient cycles, leading to less fertile soils that may hinder forest regeneration and resilience against environmental disturbances. Additionally, as tropical rainforests are critical global carbon sinks, reduced decomposition rates could impair their ability to capture and store carbon, affecting efforts to mitigate climate change.
8. Climate-Land Use Relationship: Climate change influences patterns of land use, affecting both deforestation rates and forest regeneration dynamics in a cyclical relationship where each factor influences the other.
5.2 Hypothesis (making a calculated guess about cause and effect): The researchers hypothesized that termite and fungal species richness, composition, and functions in decaying deadwood would be lower in secondary and old-growth tropical rainforests.
5.3Experiment (establishing a system to collect data to evaluate if the guess of cause and effect is factually true or not)
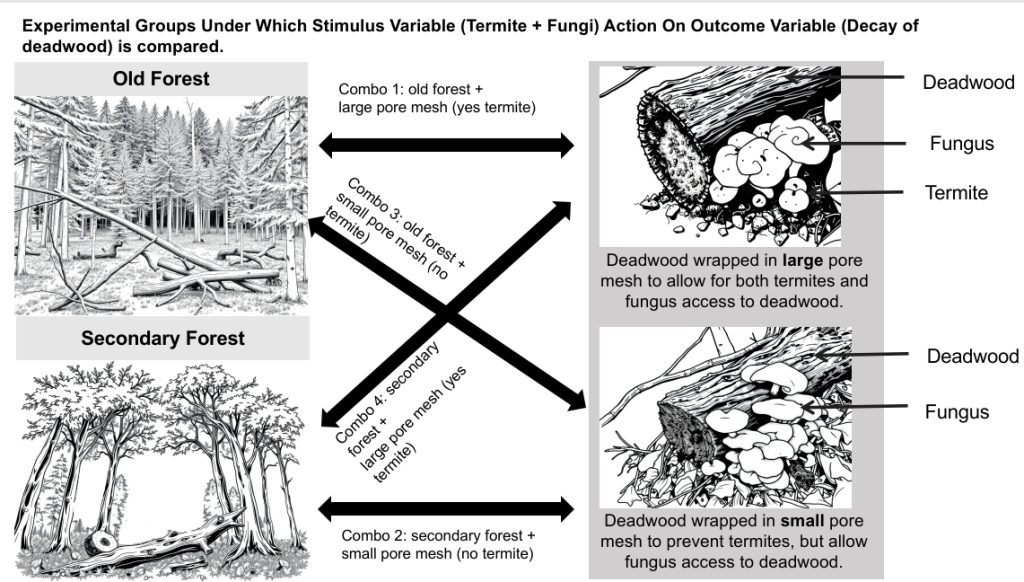
Experiments are done to examine the validity of a hypothesis and study cause and effect relationships between stimulus or independent variable and outcome or dependent variable, within the groups of focus, by observing how changing stimulus variable changes outcome variable in the groups of focus. Here stimulus or independent variables are termites and fungus, and dependent or outcome variable is deadwood decay, in the groups of focus old forest and secondary forests . As the independent or stimulus and dependent or outcome variable being studied, could also be potentially influenced by other variable called confounding or extraneous variables, which are not focus of the study, scientists hold these other variables constant, or do the experiments at a known value of the confounding or extraneous variables. Here the extraneous or confounding variables are moisture, location (latitude and longitude), decay time (duration of action of termites and fungus on deadwood). Experiment, can be broadly be divided into two parts, method and results, and thus scientific papers always have a ‘Methods’ section and a ‘Results’ section.
5.4 Experimental Methods(experimental setup and data collection from it): The researchers placed non-native pine blocks, half of which were accessible to termites, in an old-growth rainforest site as a reference and two secondary growth rainforest sites restored 4 and 8 years before the experiment. The blocks were harvested every 6 months for 4 years, and fungal ITS amplicon sequencing was used to determine wood-dwelling fungal community composition. They then collected samples from these blocks every 6 months for 4 years using mesh bags that allowed access by macroinvertebrates such as termites. In the laboratory, they examined each block to determine whether it had been discovered by termites based on signs of termite activity such as imported soil or carton. They also used DNA sequencing to confirm the presence of termites in some blocks.
5.5 Experimental Results (results are obtained by doing analysis on the data):
To analyze how systems degrade or fail over time, researchers employed the Weibull function, a versatile statistical model adept at modeling various failure patterns. Decay—or the process of progressive deterioration—can manifest in different ways, much like the aging of everyday objects. For instance, some components wear out gradually, akin to a car’s brake pads thinning with use until they finally fail. Others experience sudden, unpredictable breakdowns, resembling a lightbulb burning out without warning. The Weibull function is particularly valuable because it accommodates both types of failure patterns, whether the decline is slow and predictable or abrupt and random. By applying this model, researchers can better predict failure timelines, optimize maintenance schedules, and enhance system reliability in fields ranging from engineering to healthcare.
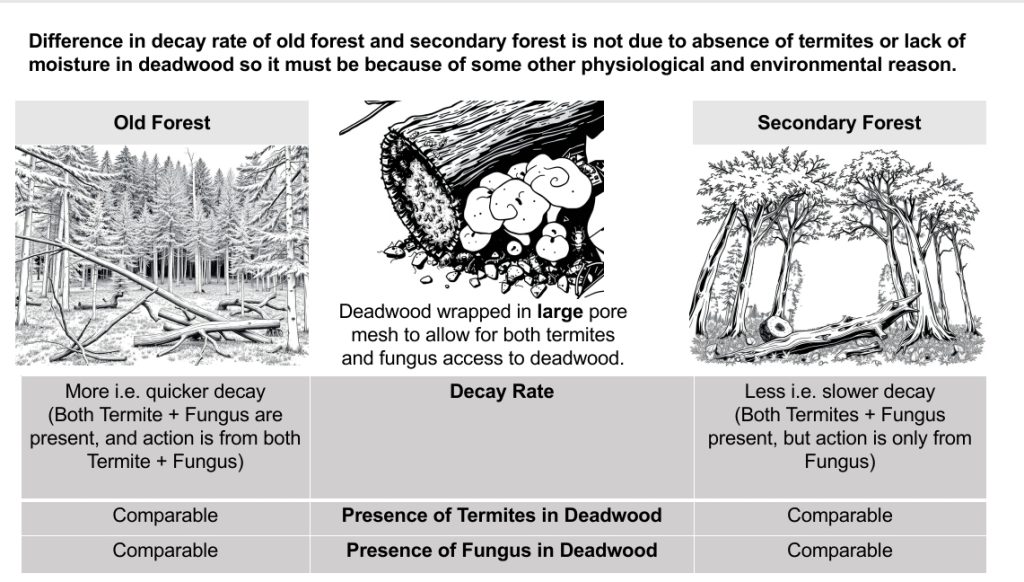
- Termites discovered similar proportions of deadwood across the secondary and old-growth rainforest sites, but the decay rates of the discovered deadwood were lower in the secondary growth rainforest.
- Fungal decay was similar to old-growth rainforest levels in the older secondary growth rainforest, but slower in the younger secondary growth rainforest, although the differences among sites were small.
- Wood-dwelling fungal communities were similar between secondary and old-growth rainforests.
5.6Inference (taking results into account, hypothesis is accepted or rejected):
Contrary to common assumptions, fungal communities and their wood decay functions are resilient and recover relatively quickly within secondary growth rainforests, but termites do not, which could reduce carbon and nutrient cycling in these ecosystems. Active management methods, such as the local transplant of termite- and fungi-occupied logs, could accelerate the recovery of these ecosystems. They only provide initial insight into the recovery of fungal and termite functioning in deadwood decay after rainforest regeneration without addressing the consequences for the transfer of carbon into soil and atmosphere across global ecosystems. Inference is the crucial section of scientific papers where researchers analyze and interpret their findings to determine whether they support or refute their hypothesis. This discussion also involves exploring how the results contribute to the existing knowledge in the field, highlighting potential applications and impacts, acknowledging limitations and proposing ways to address them, and raising new questions that future studies can explore.
6. METHODS
6.1 Study Overview
The study was conducted in the lowland tropical rainforests of the Daintree region in far North Queensland, Australia. Researchers utilized two old growth rainforest sites and two secondary growth rainforest sites for this research. The old growth rainforest site had no history of clearing, while the secondary growth rainforests were replanted in 2010 and 2014.
6.2 Canopy Structure Differences
In the Daintree region, there are notable differences in canopy structure between old growth and secondary (younger) growth forests, particularly in terms of openness and height. Old growth forests typically feature taller, more vertically stratified canopies with greater structural complexity. Canopy trees in old growth forests can exceed 40 meters in height, with some conifers reaching over 64 meters tall. This results in distinct two-tiered canopies, comprising a well-developed upper conifer layer and a lower hardwood layer primarily under 32 meters tall. In contrast, secondary growth forests generally have shorter canopies, usually under 40 meters, with a denser and more uniform vertical structure lacking clear tiers.
6.3 Canopy Openness
Old growth forests are characterized by greater canopy openness, especially in the upper layers. This openness is the result of natural thinning, tree mortality, and the formation of gaps caused by branch or tree falls, all of which are crucial for maintaining biodiversity and promoting regeneration. Conversely, secondary forests tend to have denser, more closed canopies with less vertical variation in openness. The canopy in these younger forests is more uniform, exhibiting fewer large gaps compared to their old growth counterparts.
6.4 Diversity and Complexity
Diversity and structural complexity also differ significantly between the two forest types. Old growth canopies boast a higher diversity of tree species distributed throughout the canopy depth, thereby contributing to varied light environments and complex structures. In contrast, secondary forests are dominated by fast-growing, light-demanding species, resulting in lower species diversity and a simpler canopy architecture.
6.5 Changes by 2018
By 2018, canopy cover had increased in both secondary growth rainforests, accompanied by rises in litter and log/branch cover. Notably, naturally recruiting trees were able to establish themselves successfully from the surrounding rainforest into the planted matrix of both secondary growth rainforests, indicating positive ecological dynamics within these replanted areas.
6.6 Termite and Fungal Community Study
The researchers explored termite communities by collecting samples from the forest floor and analyzing their gut microbiomes alongside non-termite individuals. They examined the abundance of specific genes linked to termite metabolism, while for fungal communities, they collected sawdust samples and analyzed their composition using next-generation sequencing technology.
6.7 Methodological Differences
The study employed different methodologies for termites and fungi. Termite gut samples were taken directly from individual termites, whereas sawdust was collected from specific forest locations. While gene sequencing was used for termite analysis, next-generation sequencing was applied for fungi.
6.8 Decay Process Recovery Assessment
To assess decay processes, the researchers used wood blocks from the non-native Pinus radiata species, dried at 120°C for 48 hours to remove deterrent compounds. These blocks were wrapped in Lumite mesh to allow access to fungi but not to most macroinvertebrates, following established protocols.
6.9 Experimental Setup
A total of 240 wood blocks, split evenly between accessible and inaccessible to termites, were placed across three sites, with blocks spaced 5 meters apart. Over four years, one block from each treatment was randomly collected at the end of each dry and wet season.
6.10 Collection and Analysis of Wood Blocks
After collection, blocks were processed in the lab within 24 hours. Any signs of termite activity, such as soil or tunnels, indicated that the block had been discovered by termites. The blocks were then oven-dried at 105°C until a constant mass was achieved.
6.11 Fungal Community Analysis
Sawdust samples were collected from the wood blocks and stored in CTAB buffer for DNA extraction. This DNA was sequenced using the Illumina MiSeq platform. The resulting sequences were analyzed to identify fungal operational taxonomic units (OTUs) by comparing them to a reference database. This method allowed the researchers to classify different types of fungi present in the samples without necessarily identifying the exact species.
7. RESULTS
7.1 Figure 1 of paper: Decay of Termite-Discovered Deadwood in Secondary vs. Old Growth Rainforests
The authors’ research demonstrated that deadwood discovered by termites decayed significantly faster in old growth rainforests compared to secondary growth rainforests. They observed that undiscovered deadwood primarily underwent fungal decay pathways and decayed more slowly in the secondary rainforest from 2014 than in the old growth rainforest. However, the decay rates of undiscovered deadwood in the secondary growth rainforest from 2010 were similar to those in the old growth rainforest. These results indicate that termites play a crucial role in the decay process of deadwood. In old growth rainforests, termite-discovered deadwood decays faster than undiscovered deadwood. In contrast, decay rates in secondary growth rainforests are lower than those in old growth forests but still higher than in undiscovered deadwood. This suggests that termites consume fewer resources they encounter in secondary growth rainforests compared to those in old growth rainforests. Additionally, fungal communities differ between secondary and old growth rainforests, with fungi in the 2014 secondary forest exhibiting slower decay rates compared to those in the old growth forest. Overall, these findings highlight the importance of both termites and fungi in the decomposition process and their impact on nutrient cycling in different types of forests.
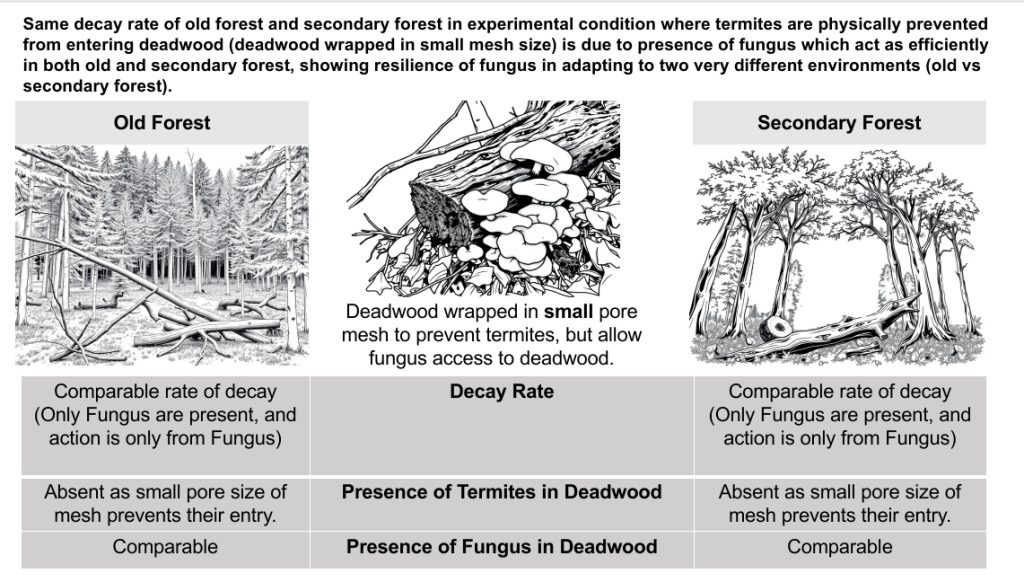
7.2Figure 2 of Paper: Fungal Decay of Deadwood and Community Composition
The authors found that undiscovered deadwood primarily underwent fungal decay pathways, which were slower in the 2014 secondary rainforest compared to the old growth rainforest. At the time of fungal community sampling (harvest 7, after 3.5 years of decay), there were no observable differences among sites in terms of mass remaining or moisture content for the deadwood that was undiscovered by termites. They also noted that the relative abundance of Agaricomycetes—a fungal group commonly involved in wood decomposition—was similar across all sites examined. Additionally, no significant differences were found in the operational taxonomic unit (OTU) richness of Agaricomycetes among sites or between those with and without termite activity.
8. DISCUSSION
8.1 Decay Processes in Rainforest Ecosystems
The study explored the decay processes of deadwood in both old-growth and secondary rainforests, emphasizing the roles of termites and fungi. Researchers monitored wooden blocks over time to compare decay rates between the two forest types. They discovered that the decay of undiscovered deadwood occurs more slowly in secondary forests, indicating higher fungal activity compared to old-growth forests. In contrast, termites exhibited a faster consumption rate, utilizing additional workers when they discovered resources.
8.2 Interactions Between Termites and Fungi
To investigate the potential symbiotic relationship between fungi and termites, researchers included wooden blocks made from the non-native species Pinus radiata during a four- to eight-year secondary growth period following colonization by Microcerotermes termites. Findings revealed that while termites decomposed deadwood at a slower rate in secondary forests, fungi maintained consistent decay rates across both forest types. Fungi typically require the mechanical breakdown provided by termites to access nutrients in deadwood, while also producing specialized enzymes for direct decomposition. This interplay accounts for variations in decay rates.
8.3 Nutrient Recycling and Ecosystem Health
Termites play a crucial role in nutrient recycling by breaking down deadwood, which nourishes other organisms and aids in carbon sequestration. However, aggressive feeding behaviors may deplete resources, negatively affecting other species. Understanding decay rates in secondary versus old-growth forests is vital for effective forest management and conservation strategies. Protecting areas rich in deadwood helps sustain diverse decomposer communities and enhances nutrient cycling, while minimizing wood waste from human activities can aid ecosystem health by reducing competition among termites.
8.4 The Ecological Significance of Deadwood
Termites create microhabitats through their decomposition activities, positively influencing various organisms and potentially altering community composition over time. The ecological significance of deadwood underscores the need for informed forest management. Its removal from managed forests can disrupt habitats and undermine nutrient cycling, highlighting the necessity to conserve deadwood in order to enhance biodiversity and maintain ecosystem integrity. Old-growth forests, abundant in deadwood, support complex habitats and diverse biological communities, unlike secondary forests that often lack this important resource.
8.5 Role of Fungi and Statistical Analysis in Decay Rates
The authors noted that undiscovered deadwood primarily undergoes fungal decay, which is slower in secondary growth forests. They found no significant differences in mass or moisture content of deadwood between sites with and without termite activity. Their aim was to elucidate the decay rates of fungi and termites in tropical rainforests, particularly focusing on secondary growth as of 2014. Conclusions indicated that fungi predominantly drive the decay process over termites, with evidence showing similar T50 values—time required for 50% mass loss—in wood accessible to termites compared to wood not exposed. The slower decay rates in younger secondary forests suggest stronger fungal dominance, and to analyze decay over time, researchers applied the Weibull function, a statistical model useful for understanding various failure patterns.
8.6 Implications of Disturbances on Decay Processes
The Weibull model employs two parameters: the shape parameter (β, beta) and the scale parameter (η, eta). The shape parameter β indicates the failure rate pattern, where β < 1 signifies decreasing failure rates, β = 1 represents a constant failure rate, and β > 1 indicates increasing failure rates typical of wear-out failures. The scale parameter complements this analysis by defining the timeframe for decay events. Overall, both termites and fungi are integral to the ecosystems of tropical rainforests, but their contributions vary depending on forest age and environmental conditions. Disturbances, such as logging or land clearing, can disrupt these decay processes, making it crucial to understand how these organisms rebound to assess ecosystem recovery over time. Just as a factory relies on a well-functioning team to operate efficiently, the balance between fungi and termites is essential for maintaining the health of rainforest ecosystems.
8.7 Exploring the Influence of Evolutionary History on Deadwood Decay Across Global Forest Ecosystems
Given Australia’s unique evolutionary history—shaped by its long separation from other landmasses—the flora and fauna of its tropical forests, such as Daintree, are distinct from those of regions like China’s Xishuangbanna, which also features tropical forests but with a different evolutionary background. Comparing deadwood decay processes across these ecosystems offers fascinating insights into how evolutionary history influences decomposition. While extending the comparison to European and Canadian forests, such as boreal and temperate forests, may introduce additional variables, it remains valuable for understanding broader ecological patterns and the effects of climate, species composition, and evolutionary history on deadwood decay.
8.8 Alexander von Humboldt and The Ecological Network: From Microbial Decomposers to Global Carbon Cycles
Both termites and fungi serve as crucial agents in tropical rainforest ecosystems, functioning as essential components in the global carbon cycle. Their contributions to decomposition and nutrient cycling vary based on forest age, successional stage, and environmental conditions. When disturbances like logging or land clearing occur, these delicate decay processes become disrupted, potentially altering carbon emissions and sequestration patterns within these vital carbon sinks. Alexander von Humboldt, the pioneering 18th-century botanist, provided the scientific foundation for understanding such ecological relationships. During his expeditions across South America for the Spanish Empire, Humboldt developed classification systems based on climate and vegetation that remain fundamental to modern ecology. His holistic approach to studying nature revealed how organisms interact with their environment and each other—insights directly applicable to contemporary research on decomposition processes involving fungi and termites. Just as Humboldt observed the interconnectedness between climate zones and biological communities, today’s ecologists recognize how the balance between fungi and termites maintains rainforest health through their complementary roles in carbon processing. Humboldt’s meticulous documentation of plant and animal distributions created an invaluable baseline for understanding ecosystem function and biodiversity patterns. This historical perspective enriches our current understanding of the complex relationships within the carbon cycle—where humans, animals, plants, and microorganisms collectively influence global carbon dynamics and, consequently, Earth’s climate stability.
REFERENCES:
[1] Friedlingstein, P., O’Sullivan, M., Jones, M. W., Andrew, R. M., Bakker, D. C. E., Birch, C. E., & Zaehle, S. (2023). Global Carbon Budget 2023. Earth System Science Data, 15(12), 5301-5369. [Global Carbon Budget 2023]
[2] Meng, W.-j.; Wen, Z.-l.; Kasanen, R.; Sun, H. and Asiegbu, F. O. (2025). Microbial communities in the phyllosphere and endosphere of Norway spruce under attack by Heterobasidion, Frontiers in Microbiology 15.
[3] Tuo, B.; Lin, L.; van Rantwijk, R. S.; van Logtestijn, R. S.; Goudzwaard, L.; Scheffers, K.; Berg, M. P.; Hefting, M. M.; Sass-Klaassen, U.; Zuo, J.; Guo, C. and Cornelissen, J. H. (2025). Positive feedback from woodpeckers on deadwood decomposition via invertebrates, Current Biology .
[4] Balonov, M. I.; Krisyuk, E. M. and Ramel, C. (1999). Environmental radioactivity, population exposure and related health risks in the east Baltic region., Scandinavian journal of work, environment & health 25 Suppl 3 : 17-32.
[5] Holl, K. D. and Kappelle, M. (1999). Tropical forest recovery and restoration, Trends in Ecology & Evolution 14 : 378-379.
[6] Neale, D. B. and Savolainen, O. (2004). Association genetics of complex traits in conifers, Trends in Plant Science 9 : 325-330.
[7] Neale, D. B. (2007). Genomics to tree breeding and forest health, Current Opinion in Genetics & Development 17 : 539-544.
[8] Edwards, D. P.; Tobias, J. A.; Sheil, D.; Meijaard, E. and Laurance, W. F. (2014). Maintaining ecosystem function and services in logged tropical forests, Trends in Ecology & Evolution 29 : 511-520.
[9] Brunet‐Navarro, P.; Jochheim, H. and Muys, B. (2016). Modelling carbon stocks and fluxes in the wood product sector: a comparative review, Global Change Biology 22 : 2555-2569.
[10] Ulyshen, M. D.; Diehl, S. V. and Jeremic, D. (2016). Termites and flooding affect microbial communities in decomposing wood, International Biodeterioration & Biodegradation 115 : 83-89.
[11] Celedon, J. M. and Bohlmann, J. (2017). An extended model of heartwood secondary metabolism informed by functional genomics, Tree Physiology 38 : 311-319.
[12] Wuyts, B.; Champneys, A. R. and House, J. I. (2017). Amazonian forest-savanna bistability and human impact, Nature Communications 8.
[13] Kerdraon, D.; Drewer, J.; Chung, A. Y. C.; Majalap, N.; Slade, E. M.; Bréchet, L.; Wallwork, A.; Castro-Trujillo, B. and Sayer, E. J. (2020). Litter Inputs, but Not Litter Diversity, Maintain Soil Processes in Degraded Tropical Forests—A Cross-Continental Comparison, Frontiers in Forests and Global Change 2.
[14] Scheidel, A. and Gingrich, S. (2020). Toward sustainable and just forest recovery: research gaps and potentials for knowledge integration, One Earth 3 : 680-690.
[15] Venäläinen, A.; Lehtonen, I.; Laapas, M.; Ruosteenoja, K.; Tikkanen, O.; Viiri, H.; Ikonen, V. and Peltola, H. (2020). Climate change induces multiple risks to boreal forests and forestry in Finland: A literature review, Global Change Biology 26 : 4178-4196.
[16] Forster, E. J.; Healey, J. R.; Dymond, C. and Styles, D. (2021). Commercial afforestation can deliver effective climate change mitigation under multiple decarbonisation pathways, Nature Communications 12.
[17] Nunes, M. H.; Jucker, T.; Riutta, T.; Svátek, M.; Kvasnica, J.; Rejžek, M.; Matula, R.; Majalap, N.; Ewers, R. M.; Swinfield, T.; Valbuena, R.; Vaughn, N. R.; Asner, G. P. and Coomes, D. A. (2021). Recovery of logged forest fragments in a human-modified tropical landscape during the 2015-16 El Niño, Nature Communications 12.
[18] Ward, E. B.; Duguid, M. C.; Kuebbing, S. E.; Lendemer, J. C. and Bradford, M. A. (2022). The functional role of ericoid mycorrhizal plants and fungi on carbon and nitrogen dynamics in forests, New Phytologist 235 : 1701-1718.
[19] Komorowicz, M.; Janiszewska-Latterini, D.; Przybylska-Balcerek, A. and Stuper-Szablewska, K. (2023). Fungal Biotransformation of Hazardous Organic Compounds in Wood Waste, Molecules 28 : 4823.
[20] May, D.; Moos, C.; Dorren, L.; Noyer, E.; Temperli, C. and Schwarz, M. (2023). Quantifying the long-term recovery of the protective effect of forests against rockfall after stand-replacing disturbances, Frontiers in Forests and Global Change 6.
[21] Fahey, T. J.; Heinz, A. K.; Mathisson, R.; Fahey, C. and Yavitt, J. B. (2024). How Much Soil Carbon is Derived from Woody Detritus? A Ten-Year Study of 13C Incorporation into Soil Organic Matter, Ecosystems 27 : 867-878.
[22] Huang, H.; Zhou, Y.; Kang, X.; Zhu, X. and Tong, X. (2024). Editorial: Water and carbon dynamics, ecosystem stability of forest and grassland in response to climate change, Frontiers in Plant Science 15.
[23] Lan, K.; Zhang, B.; Lee, T. and Yao, Y. (2024). Soil organic carbon change can reduce the climate benefits of biofuel produced from forest residues, Joule 8 : 430-449.
[24] Liu, M.; Wang, Y.; Zhang, H.; Hao, Y.; Wu, H.; Shen, H. and Zhang, P. (2024). Mechanisms of photoprotection in overwintering evergreen conifers: Sustained quenching of chlorophyll fluorescence, Plant Physiology and Biochemistry 210 : 108638.
[25] Werden, L. K.; Cole, R. J.; Schönhofer, K.; Holl, K. D.; Zahawi, R. A.; Averill, C.; Schweizer, D.; Calvo-Alvarado, J. C.; Hamilton, D.; Joyce, F. H.; San-José, M.; Hofhansl, F.; Briggs, L.; Rodríguez, D.; Tingle, J. W.; Chiriboga, F.; Broadbent, E. N.; Quirós-Cedeño, G. J. and Crowther, T. W. (2024). Assessing innovations for upscaling forest landscape restoration, One Earth 7 : 1515-1528.
[26] Wijas, B. J.; Allison, S. D.; Austin, A. T.; Cornwell, W. K.; Cornelissen, J. H. C.; Eggleton, P.; Fraver, S.; Ooi, M. K.; Powell, J. R.; Woodall, C. W. and Zanne, A. E. (2024). The Role of Deadwood in the Carbon Cycle: Implications for Models, Forest Management, and Future Climates, Annual Review of Ecology, Evolution, and Systematics 55 : 133-155.
[27] Xu, X.; Li, Y.; Li, S. and Fan, H. (2024). Post-fire forest recovery trajectory characterized by a modified LandTrendr recovery detection method: A case study of Pinus yunnanensis forests, Agricultural and Forest Meteorology 354 : 110084.
[28] Lange, H. and Hauhs, M. (2025). Complexity Analysis of Environmental Time Series, Entropy 27 : 381.
[30] The Invention Of Naure By Andrea Wulf, Published by Hodder. Royal Society Trivedi Science Book Prize Winner 2016 https://royalsociety.org/medals-and-prizes/science-book-prize/books/2016/invention-of-nature/
[31] Views of nature: or Contemplations on the sublime phenomena of creation by Alexander Von Humboldt English Translator H.G. Bohn, 1850